都市における進化を取り扱った本稿。前編では、比較的サイズの大きな集団で起こる「適応進化」を詳説しました。一方、比較的サイズの小さな集団には、また別のメカニズムで進化が起きているようです。
偶然による進化
集団中に遺伝的変異があり、特定の変異が有利/不利になるような自然選択が働けば、集団中の性質が環境に合うように変化していく――これが自然選択による進化=適応進化です。都市で起きるさまざまな生物の適応進化の事例をみると、適応進化の普遍性に感心します。そこには、偶然が入り込む隙なんてないように見えるかもしれません。確かに、多くの個体からなる集団ではその通りです。しかし実は、偶然が都市の一部の生物の運命に重大な影響を及ぼすことがあります。都市に残された自然は断片化し、孤立化しています。そのように外部から遺伝子が流入しにくい小さな集団では、偶然の遺伝子頻度の変化が、進化と言えるほどの変化となりうるのです。
これを実感するために、次のようなモデルを考えてみましょう。1000匹からなるネズミの集団があるとします。このネズミは子だくさんで1匹あたり10匹の子供を産みます。しかし、大人になるまで生存し繁殖できるのはランダムに選ばれた10%だけです(「ランダムな10%」とは、自然選択の影響がないことを意味します)。そのため次の世代も1000匹で維持されます。このネズミは、一つの遺伝子変異で生じる形質A、Bのどちらかを持っており、最初の世代ではその存在比は1:1だとします。
1000匹のネズミの集団では10000匹の子どもが生まれ、そのうち10%、つまり1000匹が生存し繁殖できます。繁殖できるネズミのうち形質Aと形質Bの比率はランダムに決まるので、496:504だったり、530:470だったりと、偶然によって少しずつ変化します。このように遺伝子の割合が偶然でゆらゆらと変化するさまを、ドリフト(漂流、浮動)するさまに例えたことから、この現象は遺伝的浮動と言われています。この例の集団は1000匹とサイズが十分に大きいので、形質AとBの割合は世代を経ても1:1から大きくずれることはないでしょう。この条件でコンピューターシミュレーションを行うと、100世代を経ても1:1から大きくはズレないことが確かめられます(図1)。
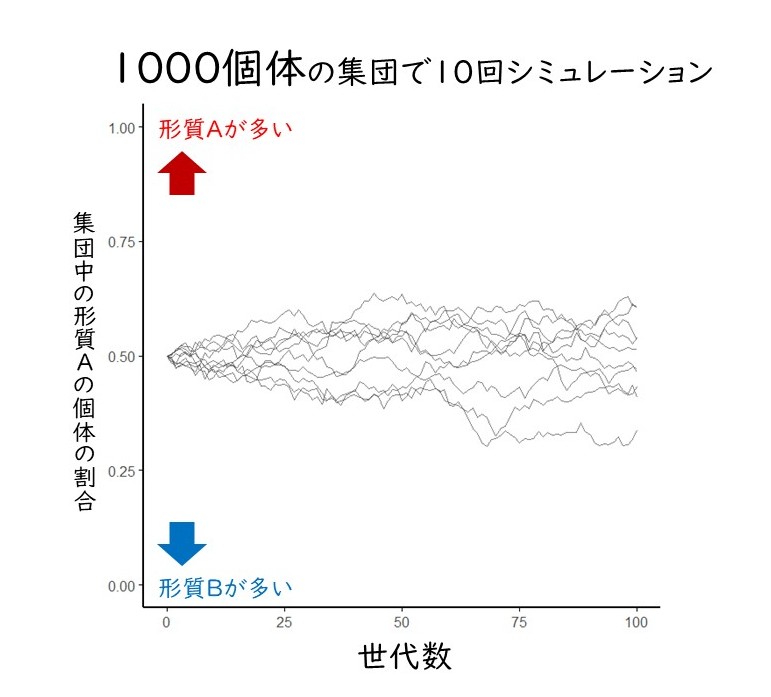
図1 形質Aが500個体、形質Bが500個体からなる1000個体の集団に遺伝的浮動が働く様子。100世代経っても、どちらかの形質だけになることはない。
では、この集団を100分の1まで一気に小さくしましょう。この小さな集団では、100個体のネズミしか生まれません。生存・繁殖できる個体はわずか10個体です。この状況では遺伝的浮動は劇的な効果を持ちます。最初の世代では、形質Aの50個体と形質Bの50個体の100個体が生まれたとしても、数世代経過するだけでAとBの割合は1:1から大きくずれていくはずです。場合によっては、偶然によって集団全体がすべて形質Aになることも起こるでしょう。そうすると、この集団は、それ以降の世代がすべて形質Aになってしまいます。自然選択が働かなくても、集団の性質が固定されてしまうのです。ふたたびシミュレーションを行うと、25世代も経るとAかBのどちらかに形質が固定されてしまいました(図2)。
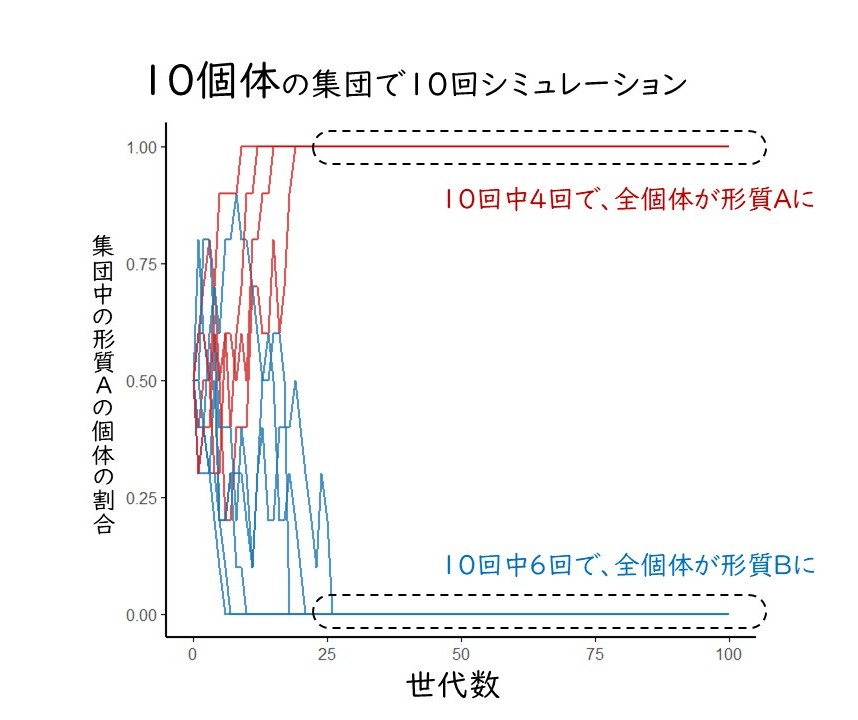
図2 形質Aが5個体、形質Bが5個体からなる10個体の集団に遺伝的浮動が働く様子。25世代経つ頃にはどの集団も、どちらかに形質だけを持つ集団に固定されている。ただし、どの形質で固定されるかは偶然による。
さらに、偶然のチカラが、自然選択のチカラを超えてしまう場合もあります。たとえば、この集団に何らかの環境変化が起き、形質Aの個体が形質Bの個体よりも20%生き残りやすくなるような自然選択が働いたとします。それでも、何世代か経つと、ランダムに選ばれる10個体が形質Bになることもあるでしょう。そうなると、本来は不利な性質であるはずの形質が集団中に固定していることになります。さらにシミュレーションしてみると、10回に1回は不利な形質Bが固定されました(図3)。
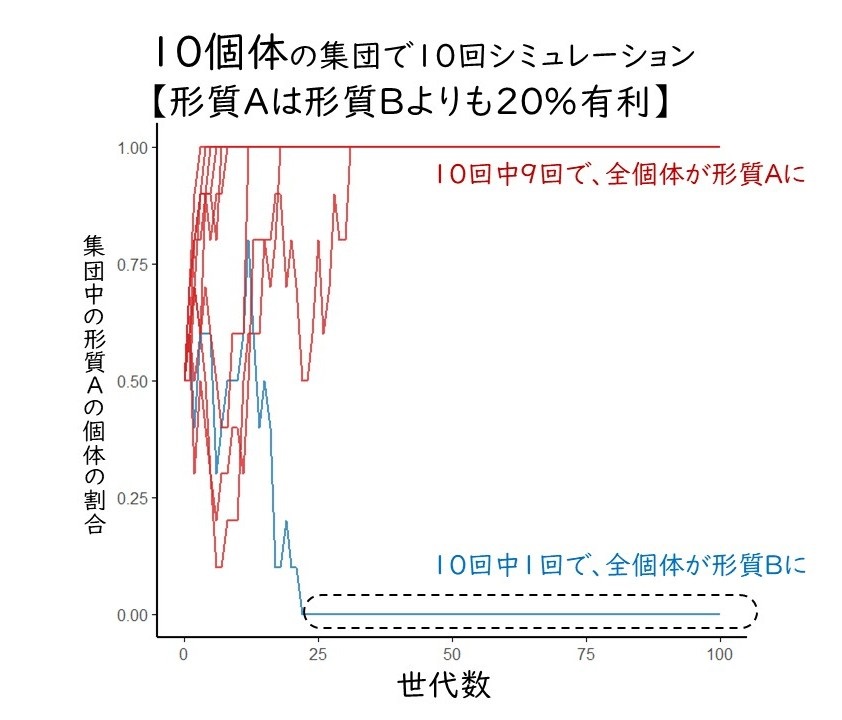
図3 形質Aが5個体、形質Bが5個体からなる10個体の集団に自然選択と遺伝的浮動が働く様子。形質Aは20%有利なはずなのに、10回中1回のシミュレーションで不利なはずの形質Bに固定される集団が出現した。
この例えでは、遺伝子は1種類で、AとBの2種類の対立遺伝子を想定しているだけですが、生物には数万種類の遺伝子があります。小さな集団で遺伝的浮動の効果が強くなると、偶然によって、それぞれの遺伝子で多様性が失われていきます。
遺伝的浮動によって遺伝的多様性が失われていることは、都市の森林や湿地など断片化され縮小した生息地を対象とした研究にも表れています。たとえば、東京西部の市街地のアカネズミを対象とした研究があります。アカネズミは野山で暮らすネズミで、都市化によって生息地が縮小している地域もあります。この研究では、孤立した生息地として国分寺市と三鷹市のアカネズミと、もう少し緑が残っていて集団間が孤立していない府中市、多摩市、昭島市、羽村市、八王子市、日野市のアカネズミの遺伝的な多様性を比較しました。その結果、遺伝的浮動のパターンから予想されるように、孤立した地域のアカネズミでは遺伝的多様性が低くなっていました(1)。同じような結果は、ニューヨークのシロアシネズミからも得られています。都市化が進んだ地域のシロアシネズミほど、遺伝的な多様性が低い傾向があったのです(2)。
近親交配によるダメージ
遺伝的浮動という偶然の進化で遺伝的多様性が低下することは、近交弱勢を介して小さな集団の存続に深刻な影響を与えます。「近交弱勢」とは、遺伝的に近縁な交配(近親交配)が続くことで、子どもの健康や生存率が低下してしまうことをいいます。私たち人間を含む生物のDNAには、個体の健康や生存に悪影響を与える有害な変異がいくつか、わずかながら存在します。しかし、個体によって、有害な変異の位置はそれぞれ異なっています。そのため、父親と母親のDNAをペアで両方持っている生物(ほとんどすべての動物・植物)は、片方の親由来のDNAに有害な変異があっても、もう片方の親由来の変異が有害でないことがほとんどです。そのため、有害な変異が実際に悪影響を及ぼすことはほとんどありません(図4)。
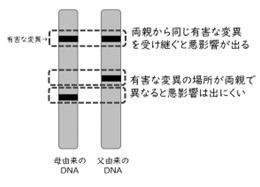
図4 有害な遺伝子が悪影響を及ぼすしくみ
ここで、近交弱勢の具体的な例として、遺伝的に近い近親者同士の交配を考えてみましょう。きょうだいやいとこなどの近親者は同じ場所に有害な変異を持っている可能性が、近親者ではない人よりも高くなります。同じ場所に有害な変異を持つ親同士から生まれた子どもは、有害な変異をペアで持ってしまい、直接的な悪影響を受けてしまいます(図5)。近親者同士の交配が続くことで、悪影響を発現する遺伝子が多くなり、子どもの健康や生存率が低下するのです。個体数が急激に減少している集団では、遺伝的な多様性が低下し、遺伝的に近い個体と交配する確率が高くなります。その結果、有害な変異の悪影響が少しずつ蓄積していき、交配しても子どもができにくく、できた子どもの生存率や生殖能力も低下する、つまり近交弱勢が生じてしまうのです。
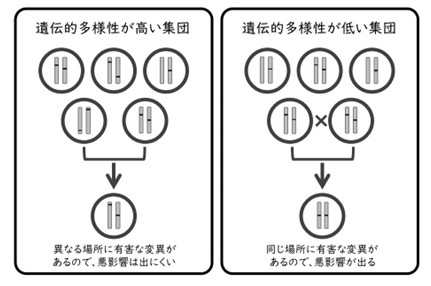
図5 遺伝的な多様性が低い集団では、近交弱勢によって遺伝的な悪影響が出やすい
北米のソウゲンライチョウ(図6)の例は、遺伝的多様性の低下のリスクを明確に示しています。ソウゲンライチョウは北米西部の広大なステップ地帯に生息していた鳥類です。イリノイ州では19世紀半ばには数百万羽がいたと推定されています。しかし、都市化や農地の拡大で草原が失われたことで、1990年にはたった数十羽にまで激減してしまいました。DNAを解析することで、個体数の減少とともに遺伝的な多様性も減少し、近親交配が起き、近交弱勢が起きていることがわかりました。最終的に、近交弱勢の影響で、卵が受精したとしても雛が孵化しないケースが多くなってしまっていました。こうなると、生息地である草原を再生しても、もはや個体数は回復しませんでした。遺伝的な多様性の低下が、絶滅リスクに直結することがわかります。

図6 ソウゲンライチョウ
こういった絶滅危惧種における遺伝的多様性の低下の事例は枚挙にいとまがありません。ただし、ソウゲンライチョウの事例は他の多くの絶滅危惧種と違って、悲しい話だけでは終わりません。イリノイ州の地元当局は、遺伝的多様性や近交弱勢の調査結果に基づいてこの地域のソウゲンライチョウ集団の絶滅は避けられないと判断し、他の州からソウゲンライチョウを移入させることを決断しました。1992年から1996年の間に、近隣のミネソタ、カンザス、ネブラスカ各州の集団から271羽が放鳥されました。放鳥後の慎重なモニタリングから、個体数が回復したこと、そして、近交弱勢がなくなったことが確認されています。卵が正常に孵化するようになったのです(3)。
ソウゲンライチョウの事例は、集団遺伝学が集団の遺伝的多様性や近親交配など絶滅リスクに関わる情報を記述するだけでなく、絶滅から救うための選択肢を提供することを示しています。ただし、上記の移入という手段が実行に移されることは、なかなか多くありません。それは、さまざまな課題やリスクがあるためです。たとえば、他集団から個体を移入し交配させることで、元の集団が持っている遺伝的な固有性が失われます。また、移入個体が新しい環境に適応できない場合もあります。これらの課題を克服するためには、移入前の詳細な調査や、地域で保全に関わる人たちとの合意形成、移入後のモニタリング、そして適切な管理計画が必要です。
進化を知って保全に貢献する
都市は、自然生態系を蚕食しながら発展してきました。都市に残された森林や湿地など貴重な自然は、断片化し、孤立化し、小さくなった生態系です。そういった生態系に棲む生物は、遺伝的浮動による遺伝的多様性の低下や近親交配などのリスクにさらされています。そのため、都市で生物の保全を行う際は、遺伝学的な知見が重要になってきます。
保全の計画を立てるには、どこに、どのように、どれだけ着手するかを決めることが大切です。調査の際、遺伝学の知見に基づいて土地の孤立性や近親交配リスクに着目してデータを収集、分析できれば、どの土地を優先して保全すればよいかを判断できます。また、孤立した生息地同士をつなぐ判断もできるようになります。他地域からの移入を考える際も、どの集団をどれだけ移入させるかを考える材料になります。生物多様性には遺伝子/種/生態系という3つの階層性があることを思い出してください(第2回参照)。最もスケールの小さい、遺伝子多様性(種内の遺伝的多様性)は、このような理由で大事なのです。
実は、遺伝的浮動という偶然による進化だけでなく、自然選択による進化を考慮することも都市の生物多様性保全にとって大事かもしれません。たとえば、ある種のネズミを考えてみましょう。都市の集団では捕食者や寄生者がいないことが選択圧となり、捕食者や寄生者に対抗するような形質が失われ脆弱になるような適応進化を遂げているかもしれません。このようなネズミ集団は、都市の自然再生によって捕食者や寄生者が増えてしまうと、絶滅リスクが上昇するでしょう。また都市の生物では、強い物理ストレスが選択圧となってさまざまなストレス耐性が進化している可能性もあります。このような適応進化は、都市という特殊な環境で生き残るために重要であり、もし周辺の集団から他個体を移入・交配させてしまうと、都市集団の生存率が低下するリスクがあるかもしれません。人間の社会経済や在来生態系に大きな悪影響を及ぼす侵略的な外来種が、都市に適応することで、より侵略的になることもあります。
都市に残された絶滅危惧種を守るためにも、都市の集団でどんな遺伝的な変化(すなわち進化)が起きているかを知ることが役に立つはずです。進化の知識は、人間が都市の多様な生物と、これからもうまく長く付き合っていくために必要不可欠です。
図版出典
図1–5 著者作成
図6 Wikimedia Commons「Greater Prairie Chicken (Tympanuchus cupido) (19729007314).jpg」より。
注
- Hirota et al., “Population structure of the large Japanese field mouse, Apodemus speciosus (Rodentia: Muridae), in suburban landscape, based on mitochondrial D-loop sequences” Molecular Ecology 13(11) 3275-82(2004).
- Jason Munshi-South, Christine P. Zolnik, Stephen E. Harris, “Population genomics of the Anthropocene: urbanization is negatively associated with genome-wide variation in white-footed mouse populations” Evolutionary Applications 9(4) 519-629(2016).
- Westemeier R. L. et al,. “Tracking the long-term decline and recovery of an isolated population.” Science 282(5394) 1695-8(1998).
過去の連載記事
(なぜ人は自然を守りたいのか?)「気持ち」でながめる自然保護(後編)
2026年3月2日
「気持ち」でながめる自然保護(前編)
2026年2月2日
農業と進化(後編)
2026年1月5日
農業と進化(前編)
2025年12月1日
農業の二面性、その複雑なメカニズム(後編)
2025年11月4日
農業の二面性、その複雑なメカニズム(前編)
2025年10月1日
都市で起きる進化(後編)
2025年9月1日
都市で起きる進化(前編)
2025年8月1日
都市の生物多様性とその恩恵
2025年6月2日
生物多様性保全のカギ② 進化生物学
2025年4月1日
生物多様性保全のカギ① 生態学
2025年2月3日
生物多様性を腑分けする
2024年12月2日