世界中で進められている生物多様性の保全や再生は、生態学や進化学と言われる生物学の一分野を学問的支柱としています。これらの学問は、生態系の中の複雑なやりとりを解きほぐし、生物多様性が生まれる歴史やそのプロセスを解明し、人間が自然から受けている恩恵を明らかにし、自然を再生・復元するための方法を検証してきました。
このように説明すると、生態学や進化学は、生物多様性を「守る」ことを目的とした学問だと思われるかもしれません。実は、それは違います。確かに生態学や進化学は生物多様性の保全や再生に不可欠ですし、生態学と進化学の知見を生物保全に直接利用することを考える、保全生態学という学問分野もあります(1)。しかし、生態学や進化学の下で行わる研究は、必ずしも保全を第一の目的としたものばかりではありません。では、生態学や進化学の中では、どんなテーマで研究が行われているでしょうか。一例として、2023年の日本生態学会の全国大会でポスター発表賞を獲得したテーマを見てみましょう(2)。
「日本産被食散布型植物の各階層における果実の色の多様性」
「積雪下で発芽することの適応的意義―展葉日と成長率に注目して―」
「シカのヌタ場が両生類の繁殖に与える影響」
「東京都都市部における社寺林の鳥類多様性維持の役割の解明」
「カニの移動方向はどのように進化したのか? 現生種情報を用いた祖先形質の復元」
これをもう少しわかりやすい言葉にすると、こうなります。「なぜ果物の色はこんなにいろいろあるのだろう?」「雪が積もっているのに発芽する植物があるのはなぜだろう?」「シカが泥浴びする場所と両生類とにどんな関係がある?」「都市の神社や寺の林は鳥の多様性と関係があるのか?」「カニはどう進化してあんなふうに動くようになったのだろう?」
これは数百題ある講演内容のほんの一部ですが、こうしてみると、必ずしも生物多様性保全のための学問ではない、というのがわかると思います。こういった多様な疑問に挑戦するのが、基礎科学としての生態学や進化学であり、いわば、生き物の不思議を解明する学問なのです。
役に立つ生態学
しかし現在、生物多様性の保全や再生を目指した世界的な取り組み(ネイチャーポジティブ)の潮流の中で、生態学や進化学の知見を持つ人たちが果たす役割は増大しています。民間企業や行政が生物多様性への取り組みを推進し、持続的に成長する未来を描くのに、不可欠なのです。ですが生態学・進化学は、「結果的に」役に立つ学問になったにすぎません。
というのも、生態学・進化学は、ほんの少し前まで民間企業や行政から必要とされることはあまりない、「面白いけど就活の役には立たない学問」の代表のような存在でした。もっというと、生態学をひとつの基盤とする自然保護活動は、しばしば企業の邪魔をする開発反対派として疎まれることさえあったはずです。しかしそういう時代であっても、研究者たちは自分自身の好奇心に従って、生物の不思議を解明することに邁進し、後進を育成し、学問を繫いできました。だからこそ、いまがあるのです。人類の知識を拡張するという目的で基礎科学・純粋科学的研究を行い、それが結果的に役に立つことがある。これは、社会における学問の役割を考えるうえで重要です。
私自身、いまでは企業や行政からたくさん相談のご連絡をいただくのですが、5年前にはこんな未来が来るとは想像もしていませんでした。ネイチャーポジティブに関する相談を受ける際、ときどき担当の方が「実は……学生の時は○○の研究をやってたんですよ」と少し気恥ずかしそうに自己紹介してくれることがあります。○○にはたとえば、魚の行動、昆虫の分類、森林の群集、などが入ります。生態学や進化学を学んで企業や行政で働いている人が、ネイチャーポジティブ対応のキーパーソンになりつつあるのです。ネイチャーポジティブに舵を切る組織のなかには、ふつう生物多様性の専門家はいませんし、生態学や進化学を学んだことがある人ですら稀ですから、「役に立たない」生態学や進化学を学んだ人が、突然、脚光を浴びるようになったのかもしれません。
これらのエピソードは、ある学問を、役に立つ⇔役に立たないという基準で測定・評価することの難しさを示しています。いますぐに役に立つ学問だけでなく、「今は」役に立たない学問をも社会として広く涵養することが、結果的に社会を強くするのだと思います。
今回と次回では、生態学や進化学の学問的枠組みを見ながら、それがどのように現代的な生物多様性保全の支柱になってきたかを説明します。まずは生態学からです。生態学はどんなふうに生き物と環境の世界を捉え、生物多様性保全に有用なのか。①対象とする生物、②生態系の階層性、③生き物の間のつながり、という3つの観点から説明します。
生態学はすべての生物を扱う
生態学は、生物と環境、生物と生物のつながりを理解しようとする学問です。この地球の生態系は無数の生物種が相互に、またそれぞれの環境と影響し合って、複雑な関係を築いています。生態学では、その複雑性をうまく説明できる一般法則を見つけようとします。でも、それだけではありません。多様性や複雑性を生み出す一般法則に注目しつつも、例外や特殊性にも光を当てます。生態系の中にいる生物たちの一般法則と、そこから外れた特殊性、この両方を愛し探求する学問です。
一般法則や概念は、ある生き物で考案されたアイデアを全く別の生き物でも検証することを積み重ねて、最終的に多くの生物に当てはまる一般的なものとされていきます。ここでは、「メタ個体群」という概念を例に、この流れを説明してみます。
メタ個体群とは、個体の移動や遺伝子のやりとりのある小さく隔離された個体群どうしをまとめて捉える概念で、広域での生物の存続を考えるのに有効なアイデアです。この概念は、数学的な理論として提案され、その後フィンランドの小さな隔離された島々に生息するチョウのデータをモデルに集中的に検証され発展しました(3)。そして、この概念はチョウだけでなく、さまざまな昆虫・哺乳類・鳥類・微生物・植物で広く検証され、その妥当性が支持されることで生態学において重要な理論となりました。
メタ個体群という概念は、生物多様性保全や再生にも大きく貢献しました。ある生物種が存続していくためには、特定の個体群の生息地を守るだけでなく、複数の個体群間が連結し個体が移動分散できることが大事だということを示しました。そして、個体群間を緩やかに連結させることの重要性は、エコロジカル・コリドー(生態的回廊)の概念を普及させる原動力となりました。全域で生物多様性を再生できない場合でも、残存する個体群を連結させるように「通り道」を配置することで広域での保全につなげようという考えです。コリドーの有効性は、コケとコケに生息する微生物群集というミニチュア生態系を対象にした興味深い実験で鮮やかに実証されています(4)。コケを小さくし孤立させるとコケ上の微生物群集の多様性が徐々に低下するのですが、コケの間にコリドーを設置すると微生物の多様性の減少が食い止められたのです。その後、多くの研究がすすめられ、今やこの考えは、都市計画事業や国土全体での保全政策などでも一般的になりました。
メタ個体群をめぐる研究の流れは、生態学と生物多様性保全の関係をあらわす代表的なものです。チョウでの研究から提案された概念が、さまざまな動植物に当てはまる一般法則となり、生物多様性を保全するうえで重要な知見となりました。こういう意味で、生態学は生物種を限定しない、すべての生物を対象とした学問なのです。
生態学にはさまざまなスケールがある――生物多様性の階層性
生態学は生物種を限定しないだけでなく、空間スケールも限定しません。実験室のシャーレで微生物同士の競争実験をする研究があるかと思えば、衛星画像から地球全体の生物多様性を推定する研究もあります。具体例をみましょう。日本生態学会が発行するEcological Researchという生態学の学術誌があります。この雑誌の2024年3月号には、オヌカトウヒレンという非常に希少なキク科植物の発芽と実生の生育に関する研究が載っています(5)。シャーレを使って発芽条件を調べるとともに、ポットで苗を栽培し成長に影響する要因を実験的に調べています。一方、同じ2024年3月号には、日本全土、宮崎から北海道まで27地点の森林を対象とした広大なスケールの論文があります(6)。この論文では、1980年から2021年までの調査地域内の一定以上の大きさのすべての樹木個体の成長を記録した膨大なデータが公開されています。1種の希少な植物に注目して栽培を行う研究と、数百種類の樹木を対象に日本全体を対象にする大規模な研究。これが同じ生態学の中で行われているのです。
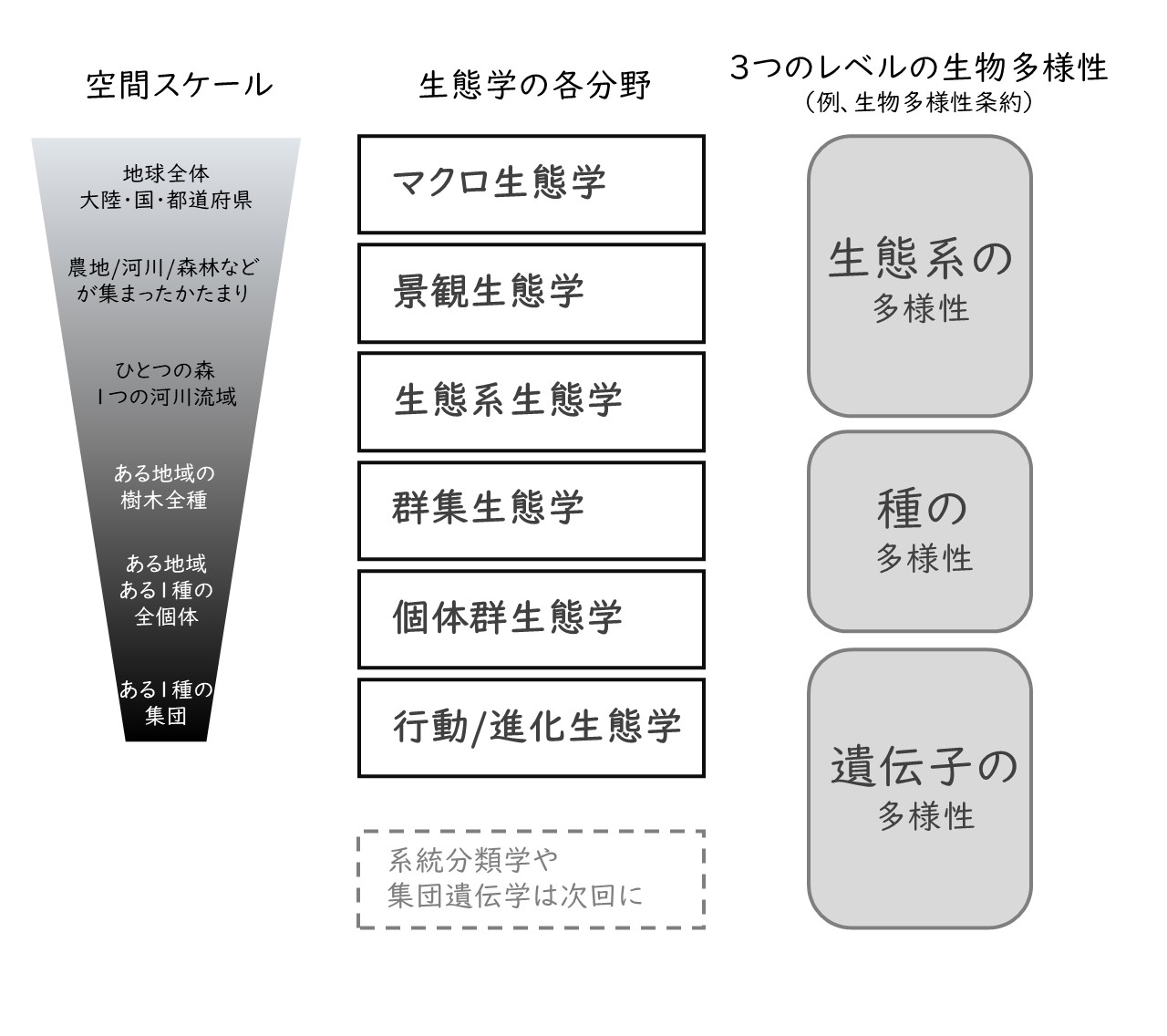
図1 生態学の分野ごとのいろいろな空間スケール
生態学が扱うスケールは、きわめて小さいものから地球規模の大きなものまでさまざまですが、扱う空間スケールに応じて分野名が付けられています(図1)。スケールの小さいところでは、特定の生物の「個体」に注目した研究分野があります。この分野は、動物の場合「行動生態学」、植物の場合「進化生態学」と言われることが多いです。特定の種の個体ごとの性質の違い(遺伝的な違いも含む)に注目して、ある環境である性質がよく観察されるのはなぜか、なぜ多様な性質が維持されているのか、などを研究します。たとえば、「なぜ都市に生えているカタバミは葉が赤いのに、農地では緑色なのか?」「なぜ動物のオスとメスの比率は多くの場合1:1なのか、しかしオシドリ、イチジクコバチ、アカシカなどそれからズレる生物が少なくないのはなぜか?」「なぜショウジョウバエには、動き回って餌を探すタイプと同じ場所にとどまるタイプが混在しているのか?」。こういった問いに挑戦します。
個体ではなく個体の集まり(個体群や集団と言います)に注目した研究は「個体群生態学」です。個体数の変動や分布などが研究されます。たとえば、ある森林に生息するシカの個体数の増減、生存率や繁殖成功、そしてそれらに影響を与える環境要因を調べる研究です。これらは絶滅危惧種の個体数を回復させるためにも、侵略的な外来種の個体数を減らすためにも必要な知見となります。また、上にあげたメタ個体群という概念は、個体群生態学の成果と言えます。
特定の種ではなく、複数の生物種に注目した研究分野は「群集生態学」と言われます。群集とはある一定の地域に住む複数の生物種の集まりのことです。群集生態学では、食う-食われるなど種間の相互作用を研究したり、複数の生物種が共存できる理由などが研究されています。食物網・食物連鎖という概念はこの分野で発展したものです。また、ある地域の群集がどれくらい多様なのかという生物多様性を測定する指標や方法論も群集生態学で発展しました。
ある地域にいる生物群集と、土壌や気候など非生物的環境との相互作用を研究する分野は「生態系生態学」です。そもそも生態学における生態系とは、生物群集とその周りの環境のことで、草原・森林・水田などが生態系の区分として用いられます。生態系生態学では、生態系を構成する生物と物理的環境に注目して研究が行われます。たとえば、生産者である植物、消費者である動物、遺骸を分解する菌類などの分解者の間で、水や太陽エネルギー、重要な元素(炭素・窒素・リンなど)がどのように移行・循環しているかが研究されてきました。これらの知見は、生物多様性保全の核となる生態系サービスの定量化につながりました。たとえば、森林生態系や湿地生態系が持つ、水を涵養したり浄化したりする能力や炭素を保持する能力は生態系生態学によって解明されてきました。
さらに広いスケールに移ります。異なる生態系がどのように組み合わさり相互に影響し合うかを研究するのが「景観生態学」です。日本の伝統的な農村の景観は、水田、森林、河川、ため池、草地など異なる生態系が組み合わさってできる里地里山と言われるものです。トンボやカエルなど、異なる生態系を行き来しないと生存できない生物がたくさんいます。また、ある生物がどれくらい遠くから移動し定着できるかは、周囲の生態系の種類や配置によって異なります。そのため、ある場所の生物多様性は、その場所の性質だけでなく、周囲の景観構造に強く影響を受けます。こういった景観生態学の知見は、広域での保全計画、都市計画、自然再生に大きく貢献しています。どこに、どのような生態系を配置すれば生物多様性を保全・再生し、生態系サービスを維持できるか、この難しい問題に挑戦するひとつの枠組みを提供してくれます。
より広域の、都道府県・国・大陸・地球全体を対象とする生態学は「マクロ生態学」と言われます。たとえば、地球全体で種の分布や多様性が偏っている理由や、そのパターンがどんな歴史的・生態学的要因で形成されたかを調べます。ここでは世界全体の衛星画像・気温・降水量・高度など大量のデータを統計的に解析するアプローチがとられます。マクロ生態学は、生態系の破壊や種の絶滅リスクパターンを明らかにすることで、人間活動が生態系に及ぼす影響を定量化してきました。またこれらの知見は、国土スケールや地球全体での保護区や優先順位の設定に使われ、生物多様性保全の役に立っています。
扱う空間スケールによる階層性は、生態学において最も重要な区分のひとつです。実際、生態学者の自己紹介は、対象とする生物・生態系と、階層性の組み合わせでなされることが多いです。たとえば、「私の専門はイノシシの個体群生態学です」「森林土壌の節足動物の群集が専門です」「海洋を対象にマクロ生態学をしています」のような感じです。ただし、少なくない生態学者が分類群や階層を越えて研究しています。私は、草本植物の進化生態学が専門ですが、群集や景観に言及した論文もあります。昆虫も対象としていましたし、動物園と協力して哺乳類の研究も行っています。
生態系の階層性を認識することは、生物多様性の保全においてもきわめて重要です。なぜなら、生物と生物、生物と環境は異なる階層で、異なる形で機能しているからです。たとえば、特定の種を保護する計画を考えましょう。そのためには、この生物の繁殖や採餌戦略を理解する(行動生態学)必要がありますし、その種の個体数の長期的な変動に与える影響(個体群生態学)や、その種の捕食者や共生者との相互作用の中での位置づけ(群集生態学)、そして生態系の物質循環の中でその種が果たす役割(生態系生態学)も考える必要があります。そして、マクロ生態学から得られるその生物の本来の生息域や環境情報は保全目標や将来計画に必須となります。このように、遺伝子/個体から地球全体まで、各階層の生態学分野が異なる視点で包括的に生物多様性保全に貢献していることがわかります。
生物多様性保全における階層性の重要性は、今に続く世界的な生物多様性保全推進のきっかけとなった1992年のリオデジャネイロで開催された地球サミットの成果からもわかります。このとき採択された「生物多様性条約」では、生物多様性には遺伝子・種・生態系という階層性があるということが言及されています(図1右)。リオ・サミットから30年以上経過し、いまや生態系の階層性は生物多様性保全の中心的な概念として普及しました。階層的なアプローチをとることで、ある種・ある個体の生存だけでなく、その種とつながりのあるさまざまな種、そしてそれらの種が生きている広域的な生態系や景観まで、さまざまなスケールで多角的に生物多様性の保全を検討できます。
生き物同士の相互作用
生態学では、生態系の中の生き物同士の相互作用(つながり)を重要視します。生態系の中で単独で生きている生物種はいません。必ず、他の種と影響し合って生きています。われわれ人間も単独では生きておらず、他の種と相互作用しながら生きています。生き物と生き物の相互作用はさまざまですが、生態学では相互作用のプラスとマイナスの組み合わせから、捕食・寄生、共生、競争の大きく3つに分類されます(図2)。相互作用する2種を考えたときに、片方の種は生態学的な利益(生存や成長、繁殖の向上)を得て、片方の種が不利益を被るような関係は「捕食」や「寄生」です。両方の種が利益を得るような関係は、「共生」と言われます。最後に、両方の種が不利益を被る関係は「競争」です。利益も不利益も起きない関係もありますが、ここでは省略します。生態系を構成する生物種は膨大なので、生物たちがどのように影響し合って生きているのか、その全貌を捉えるのは至難の業です。捕食・寄生、共生、競争の3つを区別するレンズを使うことで、複雑な生態系が整理されて見えてきます。
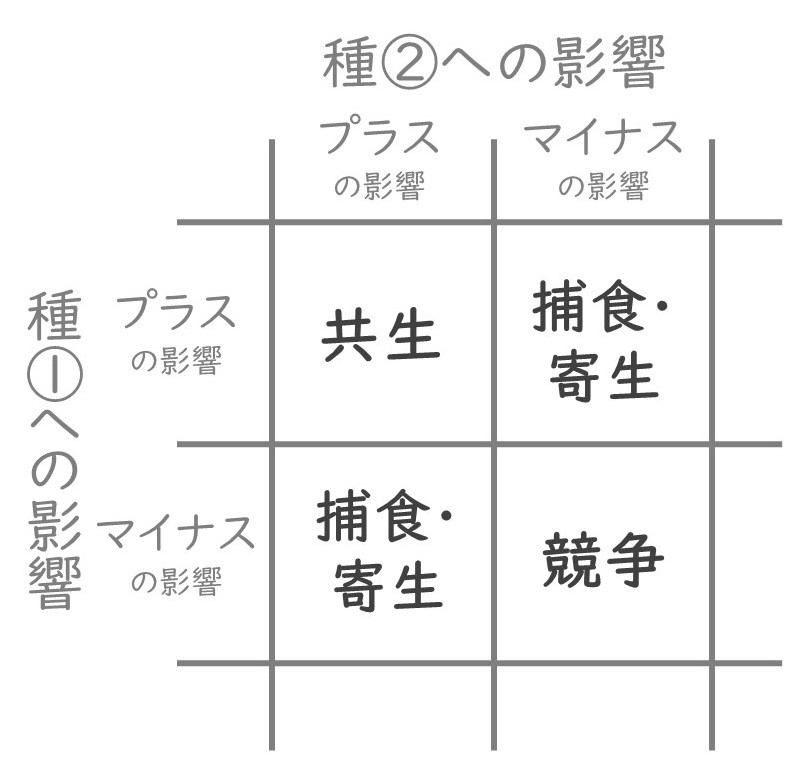
競争――資源をめぐる戦い
生態学において、競争とは、限られた資源(たとえば、食物、水、養分、生息地など)を複数の個体が共有するときに生じる相互作用のことです。競争は関わり合う両方の個体に不利益を与えます。同じ種の複数個体の場合は種内競争、異なる種の場合は種間競争と言います。競争は生態系の中で普遍的にみられる現象です。たとえば、農地生態系では、作物と雑草が光や養分をめぐって競争しています。サバンナ生態系では、シマウマやヌーやガゼルなどの大型草食動物が餌となる植物をめぐって競争しています。土壌生態系では、有機物を分解する菌類たちが炭素源(植物の遺骸など)をめぐって競争しています。競争によって個体群の成長が抑制されたり、生態系の中の種が置き換わったり、種の多様性が減少したりします。逆に、競争が、異なる種の共存を促進することもあります。
競争は生物多様性の保全を考えるうえでも重要です。侵略的な外来種は生物多様性を脅かす主因で、競争によって在来種を駆逐している例が数多く知られています。たとえば、北米原産のオオハンゴンソウは日本各地の湿地に侵入し、一面の大群落を作っています。元々生えていた在来植物は光や養分をめぐる競争によって駆逐されたと考えられています。世界中に侵入し大問題になっているヒアリ、アカカミアリ、アルゼンチンアリなども、餌をめぐる競争によって在来のアリを排除しているようです。
人間が環境変化を起こし、競争を激化させることで、競争に弱い種が消え生物多様性が低下することもあります。たとえば、農業で肥料がたくさん使われることで、近隣の草地生態系にも肥料が流れ込みます。すると、そこに生えている植物は全体として旺盛に成長し、植物間の光をめぐる競争が激化します。すると、十分な光にアクセスできない草丈が低い小さな植物種は枯れてしまいます。このような人為的な富栄養化による生物多様性の減少は、世界中の草地生態系で報告されています。こうした事態に効果的な対策を打つには、生物多様性を減少させる生物間相互作用の理解が役立ちます。この場合問題となっているのは「光をめぐる競争」の激化ですから、競争を緩和させてあげれば多様性が回復するはずです。窒素の流入を止めて富栄養化を抑制することが最もシンプルな方法ですが、生物間相互作用を考慮した解決策もあります。たとえば、ウマやヒツジなど大型の草食動物を導入して適度に背の高い植物を食べさせれば、下層に生えている植物まで光が届くようになり、小さな植物も競争に負けず存続できると考えられます。実際、このアイデアは実験的に確かめられています(7)。
捕食・寄生――網の目の関係
ある生物(捕食者)が別の生物(被食者)を消費する相互作用を捕食と言います。捕食者には利益を、被食者には不利益をもたらします。草食動物が植物を消費する「植食」も捕食のひとつです。光合成によって太陽エネルギーを有機物に変えたアブラナを、モンシロチョウの幼虫が消費します。そのモンシロチョウの幼虫を、アシナガバチが捕らえて巣に持ち帰り、アシナガバチの幼虫が消費します。アシナガバチの幼虫は、ヒメスズメバチに捕食されます。こういった捕食-被食のハシゴ上のつらなりを「食物連鎖(food chain)」と言います。ただ、捕食-被食の関係はもっと複雑で絡み合っていて、「連鎖」という一方向の表現ではちょっと足りません。なぜなら、モンシロチョウの幼虫はアシナガバチだけでなく、クモをはじめさまざまな捕食者に消費されます。そして、クモの仲間はアシナガバチを捕食するし、アシナガバチもクモの仲間を捕食します。生態学では、このような捕食-被食の複雑な絡まり合いを表現するため「食物網(food web)」という言葉もよく使われます。寄生は、ある生物(寄生者)が別の生物(宿主)から栄養を得る相互作用のことです。寄生者は宿主の体内や体表で生活し、宿主の栄養を奪いながら生活します。多くの場合、寄生は宿主を直接的な死には至らせませんが、最終的に宿主を殺すような寄生もあります(=捕食寄生)。たとえば、モンシロチョウの幼虫は、アオムシコマユバチという小さなハチによく寄生されています。このハチはモンシロチョウの幼虫に数十個の卵を産み付け、孵化したハチの幼虫は生きたアオムシの体内を食べながら成長し、最終的にアオムシの体を食い破って体外に出てそのままアオムシの上に繭を作り蛹となります。この寄生バチに寄生するハチも存在し、寄生者に寄生するこういった寄生者は、二次寄生者と言われます。スズメバチの仲間にも、スズメバチネジレバネという奇妙な寄生者がいて、働きバチのお腹の隙間で栄養を摂取しながら生活しています。寄生という生き方は奇妙にみえるかもしれませんが、実は現代人にとって最も身近な種間相互作用かもしれません。私たちの病気を引き起こすさまざまな病原菌は、人間を宿主とする寄生者です。また、シラミ、ノミ、ダニは人間や家畜の血液を餌とする外部寄生者といえます。
捕食という種間相互作用は被食者の個体数を減少させるので、生態系の動態に直接的な影響があります。たとえば、ノネコ(野生化したネコ)は多様な鳥類、爬虫類、小型哺乳類、昆虫類を捕食する非常に影響力の大きな捕食者です。ノネコは世界中で侵略的外来種として問題視されており、特に島嶼の生物多様性を大きく減少させています。たとえば、世界中の観察事例を統合した研究によると、ノネコは2,084種もの生物を捕食していて(鳥類981種、爬虫類463種、哺乳類431種、昆虫類119種、両生類57種、その他23種)、そのうち347種(16.65%)が絶滅を危惧される動物でした(8)。逆に、捕食者がいなくなることで生物多様性が減少することもあります。北米では1800~1900年初頭にかけて、入植したヨーロッパ系住民によって、ハイイロオオカミやクーガーなどの肉食哺乳類が駆除され、個体数が激減しました。この影響で増加したエルクなどの草食哺乳類が、若い樹木(ポプラやヤナギの仲間)を食べつくしてしまうことによって、樹木が大きく育たなくなってしまいました(9)。捕食者がいなくなることで植食者の個体数が増加し、植物群集を大きく変えてしまったのです。なお、一度は絶滅したハイイロオオカミを再導入したイエローストーン国立公園では、草食動物の個体数や行動範囲が大きく変化し、樹木群集が回復しつつあることが報告されています。
寄生者もまた生態系で重要な役割を果たしています。寄生生物は、宿主の神経や行動を変化させ寄生者の都合のいいように操作するという特徴があり、この特徴が時として生態系に大きな影響力を持ちます。ハリガネムシという寄生虫は、カゲロウやトビケラなど小さな水生昆虫に食べられることでその体内に入ります。成虫になった水生昆虫は上陸し、一部はカマドウマやカマキリなどの肉食性の昆虫に捕食され、それに伴ってハリガネムシもカマドウマに寄生します。カマドウマの体内でハリガネムシはどんどん大きくなると同時に、宿主の神経や行動を操りはじめ、最終的にカマドウマを川などの水辺に向かわせ、水の中に飛び込ませます。ここに至って、ハリガネムシはカマドウマのお腹から脱出し、他のハリガネムシと繁殖・産卵をするわけです。孵化した幼生はカゲロウやトビケラなどの水生昆虫に……(最初に戻る)という生き方です。見方を変えると、ハリガネムシはカマドウマを入水自殺させ、河川の渓流魚に餌を与えていることになりますが、この入水行動が生態系の食物網を大きく左右しているのです。日本の和歌山県の河川で行われた研究では、実に渓流魚の年間の摂取エネルギー量の6割近くがカマドウマから得られていることがわかっています(10)。ではもしこの重要な寄生虫がいなくなったらどうなるでしょうか。実験的にカマドウマが川に飛び込むのを抑制したところ(11)、渓流魚はカマドウマの代わりに水生昆虫をたくさん食べるようになりました。そして水生昆虫が減ることで、水中の藻類が増えたり、落葉の分解速度が遅くなるということまで観察されました(12)。たった1種の奇妙な生き方をする寄生虫が、河川の生態系全体にまで影響を与えうるのです。
共生――豊かな生態系の立役者
共生は、異なる種間で相互に利益をもたらすような生物間相互作用のことです。共生関係においては、相互作用する両者が互いに利益を得ます。たとえば、マメ科の植物は、その地下で根粒菌という細菌と共生関係にあります。根粒菌はマメ科植物の根に侵入し、植物が利用できない大気中の窒素を、植物が養分として利用できるアンモニアに変換します。対価として、植物は根粒菌に炭水化物を提供します。こうして、根粒菌は植物の成長を促し、植物は根粒菌の増殖を促しているのです。マメ科の農産物には、ダイズ(未成熟なものはエダマメ)、インゲン、アズキ、エンドウ、ラッカセイ、リョクトウ(発芽するとモヤシ)などたくさんあり、これらはすべて根粒菌と共生しています。
透き通る青い水の下の鮮やかなサンゴ礁。これも共生の産物です。サンゴはクラゲやイソギンチャクの仲間の動物で光合成ができませんが、細胞内に褐虫藻を共生させ、褐虫藻が行う光合成によって生産された糖などの有機物を受け取って生きています。逆に、サンゴは褐虫藻に住みかと栄養塩を与えます。サンゴ礁の機能は、光合成によって生態系にエネルギーを供給することだけではありません。複雑な三次元構造を持つことでさまざまな生き物に餌場や住みかを提供し、ハリケーンなどの自然災害の被害を抑制してくれます。サンゴ礁生態系は熱帯雨林と並んで、地球上で最も生物多様性が高い生態系のひとつですが、その多様性を支えるのは、共生なのです。
この地球にはさまざまな共生系がありますが、最も壮大で、最も身近な共生は、植物の繁殖をめぐる動物との共生です。植物は動けません。なので、花粉を受粉させるため、受粉してできた種子を遠くに運んでもらうため、動物のチカラを借ります。その代わりに、植物は花粉や種子を運んでくれる動物にさまざまな栄養(花の蜜や果実)を報酬として与えます。受粉をめぐる動物との共生を送粉共生、種子の散布をめぐる動物との共生を種子散布共生と言います。花は、花粉を運んでくれるハチ、チョウ、アブ、アリ、カメムシ、鳥などの動物にアピールしているのです(人間のためではありません)。また、色とりどりの美味しい果実も、果実を食べ遠く離れたところで種子を排泄してもらうためのものです(これも、人間のためではありません)。地球上の美しい色とりどりの花、芳しい香りの花をもつ植物、それにおいしい果実の数々は、植物と動物の共生関係の賜物なのです。
生態学のレンズで世界を眺めると……?
今回は、①対象とする生物、②生態系の階層性、③生き物の間のつながり、という生態学の3つの観点を説明してきました。生態学は、すべての生物を、さまざまな空間スケールから見ていて、種の存在だけでなく「つながり」からも世界を捉えようとします。図3を見てください。生態学を学ぶと、生き物たちの世界がこんなふうに見えてくる、ということを図にしてみました。
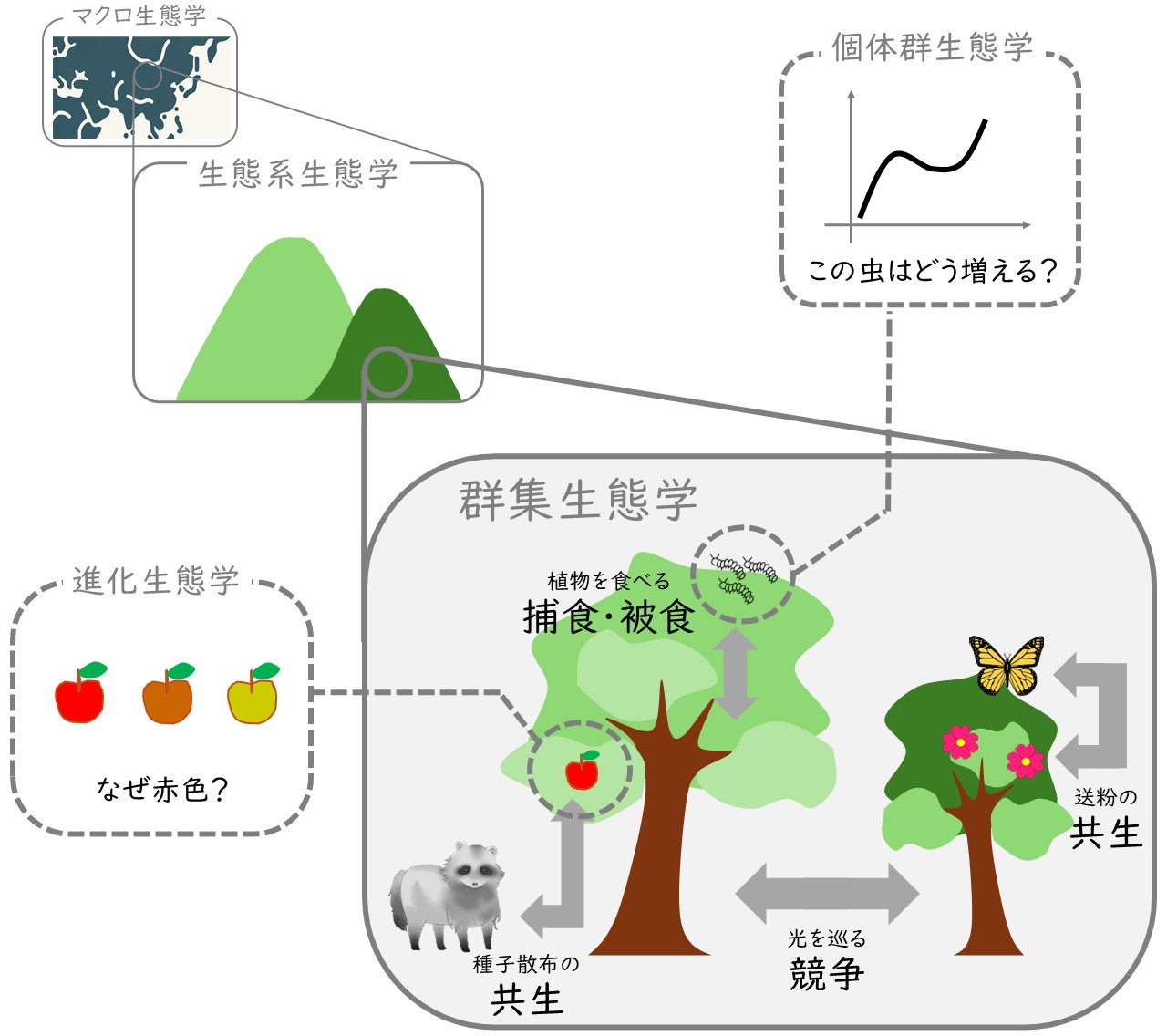
図3 自然を生態学的に捉えるということ
こういった世界の見方が、現在進められている生物多様性保全の支柱になっています。このように捉えることで、さまざまな生物が関係し合う複雑な生態系を、さまざまなスケールで理解し、適切な保全対策がとりやすくなります。生態学のレンズは生物多様性保全に必須なものです。
しかし、生態学のレンズは生物多様性保全だけに役立つのではありません。このレンズは私たちの身近な風景を少し変化させます。たとえば、道路の脇に生えている何気ない雑草たち。カタバミに黄色い花が咲いている、花粉を運ぶ送粉者はヒラタアブだろうか、カタバミだけを食べるヤマトシジミの幼虫はいるだろうか、ヤマトシジミの幼虫の捕食者はいるのだろうか。地面を歩いているのは何アリだろうか、都会のアリは何を食べているのだろうか。カタバミの横にはエノコログサ(ねこじゃらし)が生えている、エノコログサが大きくなるとカタバミは光をめぐる競争に負けるのか、エノコログサの種子は誰が散布するのだろうか、種子を食べる虫はいるのだろうか、カタバミもエノコログサも、こんな栄養のない貧弱な土壌でどうやって生きているのだろうか……などなど。このように、生態学のレンズを通してみれば道端でさえ興味をそそるフィールドになるのですから、生き物溢れる海岸・湿地・森林など、豊かな自然に足を運べば時間がいくらあってもたりないくらいです。このように、生態学のレンズは生物多様性保全に役立つだけでなく、見える世界を少しだけ楽しいものにしてくれるのです。
次回は、生物多様性保全のもう一つの柱である、進化生物学です。進化生物学のレンズは、どのように生き物と環境の世界を捉え、生物多様性保全に有用なのでしょうか。
注
- 矢原徹一・鷲谷いずみ『保全生態学入門 遺伝子からランドスケープまで (改訂版)』文一総合出版、2023年.
- 一般社団法人日本生態学会 - 第70回大会(仙台・オンライン) ポスター賞受賞者https://www.esj.ne.jp/esj/award/poster/70.html
- Ovaskainen, O., & Saastamoinen, M. “Frontiers in metapopulation biology: The legacy of Ilkka Hanski.” Annual Review of Ecology, Evolution, and Systematics, 49, 231-252 (2018).
- Gonzalez, A., Lawton, J. H., Gilbert, F. S., Blackburn, T. M., & Evans-Freke, I. "Metapopulation dynamics, abundance, and distribution in a microecosystem". Science, 281(5385), 2045-2047 (1998).
- Inoue, R., Yamamoto, A., Wasaki, J., & Nakatsubo, T., “Factors controlling germination and seedling growth of an endangered Saussurea species (Asteraceae) endemic to serpentine areas in Japan. Ecological Research, 39(4), 492-499 (2024).
- Yoshikawa, T., Totsu, K., Takeuchi, Y., Kadoya, T., Enoki, T., Fujii, S., ... & Hiura, T.,“Forest monitoring data of 45 plots across the Japanese archipelago during 1980–2021,” Ecological Research, 39(3), 391-406 (2024).
- Eskelinen, A., Harpole, W. S., Jessen, M.-T., Virtanen, R., Hautier, Y., “Light competition drives herbivore and nutrient effects on plant diversity,” Nature, 611, 301–305 (2022).
- Lepczyk, C. A., Fantle-Lepczyk, J. E., Dunham, K. D., Bonnaud, E., Lindner, J., Doherty, T. S., & Woinarski, J. C., “A global synthesis and assessment of free-ranging domestic cat diet.” Nature Communications, 14(1), 7809 (2023).
- Beschta, R. L., & Ripple, W. J., “Large predators and trophic cascades in terrestrial ecosystems of the western United States.” Biological conservation, 142(11), 2401-2414 (2009).
- Sato, T., Watanabe, K., Kanaiwa, M., Niizuma, Y., Harada, Y., & Lafferty, K. D., “Nematomorph parasites drive energy flow through a riparian ecosystem.” Ecology, 92(1), 201-207 (2011).
- この実験では高さ1.5 m、厚さ4 mmのナイロン・メッシュを川の両側に設置するとともに、そのフェンスの外(森側)に水を張ったトラップを10個設置することで、カマドウマが川に飛び込むことを防ぐ実験処理を行っています。
- Sato, T., Egusa, T., Fukushima, K., Oda, T., Ohte, N., Tokuchi, N., ... & Lafferty, K. D., “Nematomorph parasites indirectly alter the food web and ecosystem function of streams through behavioural manipulation of their cricket hosts.” Ecology Letters, 15(8), 786-793 (2012).
過去の連載記事
(なぜ人は自然を守りたいのか?)「気持ち」でながめる自然保護(後編)
2026年3月2日
「気持ち」でながめる自然保護(前編)
2026年2月2日
農業と進化(後編)
2026年1月5日
農業と進化(前編)
2025年12月1日
農業の二面性、その複雑なメカニズム(後編)
2025年11月4日
農業の二面性、その複雑なメカニズム(前編)
2025年10月1日
都市で起きる進化(後編)
2025年9月1日
都市で起きる進化(前編)
2025年8月1日
都市の生物多様性とその恩恵
2025年6月2日
生物多様性保全のカギ② 進化生物学
2025年4月1日
生物多様性保全のカギ① 生態学
2025年2月3日
生物多様性を腑分けする
2024年12月2日